


УГЛЕВОДЫ
(сахара), обширная группа полигидроксикарбо-нильных соед., входящих в состав
всех живых организмов; к У. относят также мн. производные, получаемые при хим.
Mодификации этих соед.
путем окисления, восстановления или введения разл. заместителей.
Термин "углеводы"
возник потому, что первые известные представители У. по составу отвечали ф-ле
CmH2nOn (угле-род+вода); впоследствии
были обнаружены природные У. с др. элементным составом.
Классификация и распространение.
У. принято делить на моносахариды, олигосахариды и полисахариды.
Моносахариды обычно представляют
собой полигид-роксиальдегиды (альдозы) или полигидроксикетоны (кетозы) с линейной
цепью из 3-9 атомов С, каждый из к-рых (кроме карбонильного) связан с группой
ОН. Простейший моноса-харид, глицериновый альдегид, содержит один асим.
атом С и известен в виде двух оптич. антиподов (D и L). Прочие моносахариды
имеют неск. асим. атомов С; их рассматривают как производные D- или L-глицеринового
альдегида и относят к D- или L-ряду в соответствии с абс. конфигурацией асим.
атома С, наиб. удаленного от карбонильной группы. Различия между изомерными
моносахаридами в каждом ряду обусловлены относит, конфигурацией остальных асим.
центров.
Характерное св-во моносахаридов
в р-рах - мутаротация, т. е. установление таугомерного равновесия между
ациклич. альдегидо- или кетоформой, двумя пятичленными (фураноз-ными) и двумя
шестичленными (пиранозными) полуацеталь-ными формами. Две пиранозы (как и две
фуранозы) отличаются друг от друга конфигурацией (a или b) нового асим. (аномерного)
центра, возникающего из карбонильного атома С при циклизации.
Полуацетальный (гликозидный)
гидроксил циклич. форм моносахаридов резко отличается от прочих групп ОН моно-сахарида
значительно большей склонностью к р-циям нукле-оф. замещения. Такие р-ции приводят
к образованию глико-зидов (остаток нуклеофила в гликозиде - напр. спирта.
меркаптана - носит назв. агликон). В тех случаях, когда агликоном служит др.
молекула моносахарида, образуются олиго- и полисахариды. При этом каждый остаток
моносахарида может в принципе иметь пиранозную или фуранозную форму, a- или
b-конфигурацию гликозидного центра и быть связанным с любой из гидроксильных
групп соседнего моносахарида. Поэтому число разл. по строению полимерных молекул,
к-рые теоретически можно построить даже из остатков только одного моносахарида,
представляет собой аст-рономич. величину.
К наиб, обычным и распространенным
в природе моноса-харидам относят D-глюкозу, D-галактозу, D-маннозу, D-фрук-тозу,
D-ксилозу, L-арабинозу и D-рибозу. Из представителей др. классов моносахаридов
часто встречаются: 1) дезоксиса-хара, в молекулах к-рых одна или неск.
групп ОН заменены атомами H (напр., L-рамноза, L-фукоза, 2-дезокси-D-рибоза);
2) аминосахара, где одна или неск. групп ОН заменены на аминогруппы (напр.,
2-амино-2-дезокси-D-глюкоза, или D-глюкозамин); 3) многоатомные спирты (полиолы,
альди-ты), образующиеся при восстановлении карбонильной группы моносахаридов
(D-сорбит из D-глюкозы, D-маннит из D-маннозы, и др.); 4) уроно-вые
кислоты - альдозы, у к-рых группа CH2OH окислена в карбоксильную
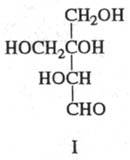
(напр., D-глюкуроновая к-та); 5) разветвленные сахара, содержащие нелинейную
цепь углеродных атомов (напр., апиоза, или 3-С- гидроксиметил-D-глицеро-тетроза;
ф-ла I); 6) высшие сахара с длиной цепи более шести атомов С (напр.,
D-седогеп-тулоза и сиаловые к-ты; ф-лы см. соотв. в статьях Пентозо-фосфатный
цикл и Моносахариды}.
За исключением D-глюкозы
и D-фруктозы своб. моносахариды встречаются в природе редко. Обычно они входят
в состав разнообразных гликозидов, олиго- и полисахаридов и м. б. получены из
них после кислотного гидролиза. Разработаны многочисл. методы хим. синтеза редких
моносахаридов исходя из более доступных.
Олигосахариды содержат
в своем составе от 2 до 10-20 моносахаридных остатков, связанных гликозидными
связями. Наиб, распространены дисахариды, выполняющие ф-цию запасных B-B: сахароза
в растениях, трегалоза в насекомых и грибах, лактоза в молоке млекопитающих.
Известны многочисл. гликозиды олигосахаридов, к к-рым относят разл. физиологически
активные в-ва, напр, гликозиды сердечные, нек-рые сапонины (в
растениях), мн. антибиотики (в грибах и бактериях), гликолипиды.
Полисахариды- высокомол.
соед., линейные или разветвленные молекулы к-рых построены из остатков моносахаридов,
связанных гликозидными связями. В состав полисахаридов могут входить также заместители
неуглеводной природы (остатки алифатич. к-т, фосфат, сульфат). В свою очередь
цепи высших олигосахаридов и полисахаридов могут присоединяться к полипептидным
цепям с образованием гликопротеинов.
Особую группу составляют
биополимеры, в молекулах к-рых остатки полиолов, гликозилполиолов, нуклеозидов
или моно- и олигосахаридов соединены не гликозидными, а фос-фодиэфирными связями.
К этой группе относят тейхоевые кислоты бактерий, компоненты клеточных
стенок нек-рых дрожжей, а также нуклеиновые кислоты, в основе к-рых лежит
поли-D-рибозофосфатная (РНК) или поли-2-дезок-си-D-рибозофосфатная (ДНК) цепь.
Физико-химические свойства.
Обилие полярных функц. групп в молекулах моносахаридов приводит к тому,
что эти в-ва легко раств. в воде и не раств. в малополярных орг. р-рителях.
Способность к таутомерным превращениям обычно затрудняет кристаллизацию моно-
и олигосахаридов, однако если такие превращения невозможны (напр., как в гликозидах
и невосстанавливающих олигосахаридах типа сахарозы), то в-ва кристаллизуются
легко. Mн. гликозиды с малополярными агликонами (напр., сапонины) проявляют
св-ва ПАВ.
Полисахариды - гидрофильные
полимеры, мн. из них образуют высоковязкие водные р-ры (растит, слизи, гиалуроно-вая
к-та; ф-лу последней см. в ст. Мукополисахариды), а в ряде случаев (в
результате своеобразной межмол. ассоциации) - прочные гели (агар, алъгиновые
кислоты, каррагинаны, пектины). Нек-рые полисахариды образуют высокоупорядоченные
надмолекулярные структуры, препятствующие гидратации отдельных молекул; такие
полисахариды (напр., хитин, целлюлоза)не раств. в воде.
Биологическая роль.
Ф-ции углеводов в живых организмах чрезвычайно многообразны. В растениях
моносахариды являются первичными продуктами фотосинтеза и служат исходными
соед. для биосинтеза гликозидов и полисахаридов, а также др. классов B-B (аминокислот,
жирных K-T, фенолов и др.). Эти превращения осуществляются ферментами, субстратами
для к-рых служат, как правило, богатые энергией фос-форилир. производные Сахаров,
гл. обр. нуклеозиддифосфат-сахара.
У. запасаются в растениях
(в виде крахмала), животных, бактериях и грибах (в виде гликогена),
где служат энергетич. резервом. Источником энергии являются р-ции расщепления
глюкозы, образующейся из этих полисахаридов, по гликоли-тич. или окислит. пути
(см. Гликолиз). В виде гликозидов в растениях и животных осуществляется
транспорт разл. метаболитов. Полисахариды и более сложные углеводсодержащие
полимеры выполняют в живых организмах опорные ф-ции. Жесткая клеточная стенка
у высших растений представляет собой сложный комплекс из целлюлозы, гемицеллюлоз
и пектинов. Армирующим полимером в клеточной стенке бактерий служат пептидогликаны
(муреины), а в клеточной стенке грибов и наружных покровах членистоногих
- хитин. В организме животных опорные ф-ции выполняют протео-гликаны соединит,
ткани, углеводная часть молекул к-рых представлена сульфатир. мукополисахаридами.
Эти в-ва участвуют в обеспечении специфич. физ.-хим. CB-B таких тканей, как
кости, хрящи, сухожилия, кожа. Будучи гидрофильными полианионами, эти полисахариды
способствуют также поддержанию водного баланса и избират. ионной проницаемости
клеток. Аналогичные ф-ции в морских многоклеточных водорослях выполняют сульфатир.
галактаны (красные водоросли) или более сложные сульфатир. гетерополисахари-ды
(бурые и зеленые водоросли); в растущих и сочных тканях высших растений эту
ф-цию выполняют пектины.
Особенно ответственна роль
сложных У. в образовании клеточных пов-стей и мембран и придании им специфич.
св-в. Так, гликолипиды - важнейшие компоненты мембран нервных клеток и оболочек
эритроцитов, а липополисахариды -наружной оболочки грамотрицат. бактерий.
У. клеточной пов-сти часто определяют специфичность иммунологич. р-ций (групповые
в-ва крови, бактериальные антигены) и взаимод. клеток с вирусами. Углеводные
структуры принимают участие и в др. высокоспецифич. явлениях клеточного взаимод.,
таких, как оплодотворение, узнавание клеток при тканевой дифференциации, отторжение
чужеродных тканей и т. д.
Практическое использование.
У. составляют главную часть пищ. рациона человека, в связи с чем широко
используются в пищ. и кондитерской пром-сти (крахмал, сахароза и др.). Кроме
того, в пищ. технологии применяют структурир. в-ва полисахаридной природы, не
имеющие сами по себе пищ. ценности,- гелеобразователи, загустители, стабилизаторы
суспензий и эмульсий (альгинаты, агар, пектины, растит. галактоманнаны и др.).
Превращения моносахаридов
при спиртовом брожении лежат в основе процессов получения этанола, пивоварения,
хлебопечения; др. виды брожения позволяют получать из Сахаров биотехнол. методами
глицерин, молочную, лимонную, глюконовую к-ты и мн. др. в-ва.
Глюкозу, аскорбиновую к-ту,
углеводсодержащие антибиотики, гепарин широко применяют в медицине. Целлюлоза
служит основой для получения вискозного волокна, бумаги, нек-рых пластмасс,
BB и др. Сахарозу и растит, полисахари-ды рассматривают как перспективное возобновляемое
сырье, способное в будущем заменить нефть в пром. орг. синтезе. Моносахариды
используют в качестве доступных хиральных исходных соед. для синтеза сложных
прир. B-B неуглеводной природы.
Лит.: Химия углеводов,
M., 1967; Общая органическая химия, пер. с англ., т. 11, M., 1986, с. 127-299.
А. И. Усов.