


ТЕЙХОЕВЫЕ КИСЛОТЫ
(от греч. teichos-стена), углеводные фосфатсодержащие полимеры бактериального
происхождения.
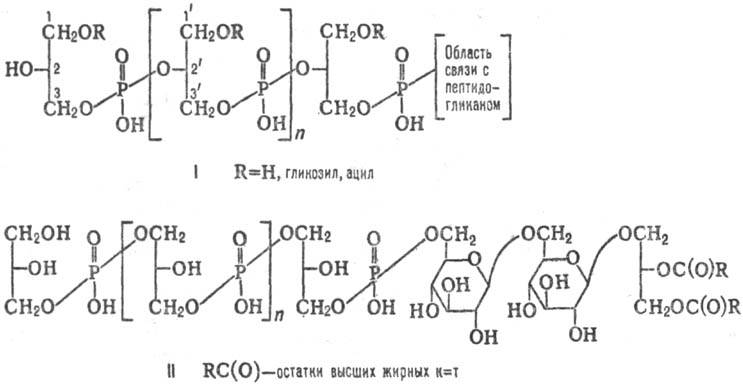
Молекулы Т.к. построены
из повторяющихся звеньев, представляющих собой полиолы или гликозилполиолы,
соединенные фосфодиэфирными связями. В зависимости от природы полиола различают
глицерин-, рибит- и ман-нит-Т. к. Фосфодиэфирные связи в поли(глицерофосфатных)
Т.к. (ф-ла I) могут соединять положения 3,2' или 1,3', а в поли(рибитфосфатных)
Т.к. установлены только связи 1,5'; строение маннит-Т.к. изучено недостаточно.
Гидрок-сильные группы полиолов могут быть гликозилированы (остатками глюкозы,
галактозы, маннозы, рамнозы, глю-козамина, галактозамина, изредка О-метилпроизводными
галактозы и рамнозы) или ацилированы (остатками D-ала-нина, L-лизина, уксусной
и янтарной к-т). В нек-рых Т.к. остатки Сахаров, гликозилирующих полиол, включены
в главную цепь полимера. В молекулах Т.к. n обычно от 10 до 40 и может
достигать 60. Сведения о вторичной структуре указывают на возможную спиральную
конфор-мацию Т.к.
Т.к. клеточных стенок (их
содержание может достигать 75% от сухой массы этих стенок) ковалентно связаны
с пептидогликанами, причем область связывания представляет собой олигомерное
звено, содержащее аминосахара и глицерофосфат, но отличающееся по строению от
повторяющихся звеньев Т.к. Как правило, ближайшим к пепти-догликану является
остаток глюкозамина; атом С-1 этого моносахарида связан фосфодиэфирной связью
с атомом С-6 остатка мурамовой к-ты пептидогликана, а с др. стороны звена (ближайшей
к Т.к.) остаток глицерофосфата фосфодиэфирной связью соединен с остатком полиола
Т.к.
Мембранные Т.к. получили назв. липотейхоевых к-т (липо-Т.к.). Они представляют собой чаще всего поли(гли-церофосфатные) цепи (фосфодиэфирная связь соединяет положения 1,3'), соединенные фосфодиэфирной связью с атомом С-6 гексопиранозильного остатка гликолипида (II). Благодаря липидной части липо-Т. к. удерживаются в мембране, а поли(глицерофосфатная) цепь располагается на ее внеш. стороне и иногда пронизывает клеточную стенку и выходит на пов-сть клетки. Содержание липо-Т. к. составляет 0,4-1,6% сухой массы клетки.
Т. к. раств. в воде, для
выделения из клеточной стенки их обычно экстрагируют 10%-ной трихлоруксусной
к-той при 2-4 °С в течение 24 ч с послед. осаждением этанолом или ацетоном.
Более продолжит. экстракция может приводить к частичному гидролизу фосфодиэфирных
связей или потере О-ацильных заместителей. Иногда Т.к. экстрагируют водным р-ром
NaOH или диметилгидразином, но в этих условиях более вероятна частичная деструкция
полимера. Липо-Т. к. выделяют обработкой разрушенных клеток водным фенолом.
Дальнейшую очистку Т.к. проводят методами гель-хроматографии, ионообменной хроматографии,
высоковольтного электрофореза на бумаге и аффинной хроматографии на лектинах.
Определение первичной структуры
Т.к. хим. методами основано на расщеплении молекул на фрагменты действием к-т
(2 н. НС1, 48% HF) или щелочей (1 н. NaOH) с послед. разделением и установлением
строения этих фрагментов. Важным физ.-хим. недеструктивным методом структурного
анализа служит ПМР и спектроскопия ЯМР 13С и 31Р.
Донорами мономерных звеньев
при биосинтезе Т. к. служат цитидиндифосфатглицерин, цитидиндифосфатрибит или
нуклеозиддифосфатсахара. Синтез цепи начинается с образования олигомерной области
на участке связи Т.к. с пептидогликаном при участии липидных переносчиков полипренольной
природы. Далее к образовавшемуся олиго-меру последовательно присоединяются полиолфосфатные
остатки от соответствующих нуклеотидных предшественников. При биосинтезе липо-Т.
к. в образовании поли(глице-рофосфатной) цепи нуклеотидный предшественник не
участвует, а донором глицерофосфатных остатков (имеющих по сравнению с ф-лой
I противоположную стереохим. конфигурацию) служит фосфатидилглицерин.
Ф-ции Т.к. в бактериальной
клетке связаны с ионным обменом и регуляцией работы автолитич. ферментов (катализируют
гидролиз сложного биополимера, составляющего каркас клеточной стенки), к-рые
активны при росте и делении клеток. Мутантные клетки бактерий, лишенные Т.к.,
оказываются нежизнеспособными. К вторичным ф-циям Т.к. относят их антигенные
св-ва и связывание фагов. Стрептококковые, стафилококковые и др. бактериальные
инфекции человека и животных сопровождаются выходом Т.к. в организм, что приводит
к развитию постинфекц. осложнений в виде эндокардитов, нефритов, артритов и
др.
Т.к. открыты в 1958 в клеточных
стенках грамположит. бактерий. Позже аналогичные полимеры были найдены в капсулах
грамотрицат. бактерий и в бактериальных мембранах.
Лит.: Вaddiley J.,
"Essays Biochem.", 1972, v. 8, p. 35-77; Naumova I. B., "Microbiological
Sciences", 1988, v. 5, № 9, p. 275-79; Fischer W., Physiology of lipoteichoic
acids in bacteria, в кн.: Advances in microbial physiology, ed. by A.H. Rose,
D.W. Tempest, v. 29, L., 1988, p. 233-302. И.Б. Наумова.