


ПРОТРОМБИНОВЫЙ КОМПЛЕКС,
группа гликопро-теинов плазмы крови, включающая факторы ее свертывания-II
(протромбин), VII, IX и X, а также регуляторные белки СИ и S, к-рые участвуют
в центр. звене каскада последоват. р-ций активации белков-предшественников,
приводящих к образованию тромбина.

Белки П. к. (см. табл.)
характеризуются гомологией первичной и третичной структур, общностью механизмов
биосинтеза и активации в соответствующие активные формы-IIа (тромбин),
VIIa, IXa, Ха и протеин СИа. Белки
П. к. синтезируются в печени и претерпевают пострибосомаль-ную модификацию в
микросомах перед секрецией в кровь. Эта модификация, контролируемая витамином
К, заключается в карбоксилировании 10-12 остатков глутаминовой к-ты в N-концевой
области каждого из белков с образованием остатков g-карбоксиглутаминовой
к-ты. Эти остатки обеспечивают способность белков П. к. связывать ионы Са2
+ , через к-рые происходит присоединение белков к отрицательно заряженной
пов-сти фосфолипидов клеточных мембран. Связывание необходимо для концентрирования
белков П.к., достижения высоких скоростей протекания последоват. р-ций каскада
и их регуляции.
Дефицит витамина К или
введение в организм его антагонистов (напр., дикумарина) приводит к снижению
или полному подавлению зависимой от этого витамина р-ции карбоксилирования остатков
глутаминовой к-ты. В результате образуются неполноценные белки П. к. и снижается
свертываемость крови. В белках П. к., за исключением протромбина, обнаружена
мало распространенная в живых организмах b-гидроксиаспарагиновая к-та,
функцион. значение к-рой не выяснено. Известна первичная структура белков П.
к., а также выделены гены, кодирующие их биосинтез.
Активные формы белков П.
к. представляют собой сери-новые протеолитич. ферменты, у к-рых каталитич. участок
активного центра расположен в С-концевых областях тяжелых цепей молекулы. После
активации проферментов (в результате элиминирования пептидных фрагментов) С-кон-цевая
область ферментов остается связанной с N-концевым доменом дисульфидными связями,
благодаря чему осуществляется оптимальная ориентация белков на мембране клетки.
Исключение-фермент тромбин, к-рый в результате активации протромбина лишается
домена, содержащего Са2+-связывающие участки.
Каталитич. св-ва ферментов
усиливаются путем их комп-лексообразования с кофакторами или регуляторными белками
на пов-сти клеточных мембран. К ним относят: фактор VIII-кофактор фактора IХа,
фактор V-кофактор фактора Ха, тканевый фактор (ТФ) - кофактор фактора
VIIa, протеин S и тромбомодулин - кофактор протеина СИа.
Регулятор-ные белки обеспечивают оптимальную локализацию ферментов вблизи соответствующих
субстратов, благодаря чему скорость активации проферментов увеличивается в десятки
тысяч раз и более.
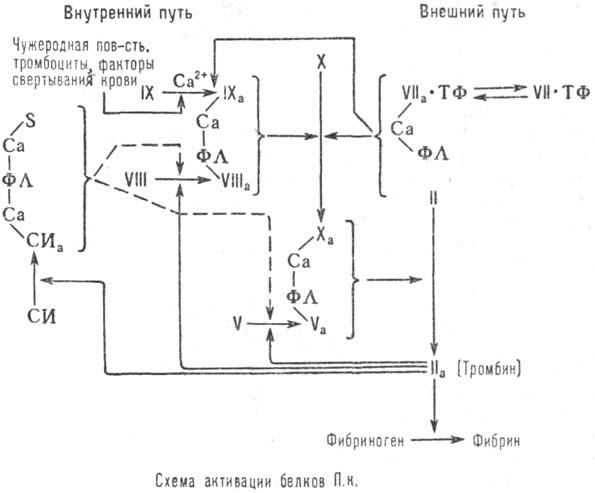
Механизм превращения белков
П. к. (см. схему) включает внеш. путь, в к-ром участвует тканевый фактор-липопро-теин,
образующийся при повреждении ткани, и внутренний. Сплошные стрелки на схеме
показывают пути активации белков П. к., пунктирные-пути ингибирования этого
процесса (ФЛ - фосфолипиды мембраны клетки). Тромбин участвует в конечной стадии
образования кровяного сгустка путем активации превращения фибриногена в фибрин.
Лит.: Страйер Л.,
Биохимия, пер. с англ., т. 1, М., 1984, с. 152-76; Jackson С, Nemerson Y., "Ann.
Rev. Biochem.", 1980, v. 49, p. 765-811.
И. П. Баскова.