


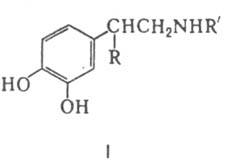
рост молочных желез, лактацию и др.). Известно также стимулирующее влияние дофамина на гликогенолиз, уровень глюкозы в крови, диурез, кровоток в почках. Нарушение синтеза дофамина в мозгу - причина возникновения болезни Паркинсона. Дофамин выделяется из пресинаптич. окончаний и действует на постсинаптич. рецепторы, к-рые либо активируют фермент адепилатцтлазу (рецепторы типа Д-1), либо оказывают на аденилатцик-лазу ингибирующее влияние и повышают уровень внутриклеточного Са2+ в результате активации фосфоинозитидного обмена (гидролиза полифосфоинозитидов до диацил-глицерина и инозитолфосфатов) или открывания рецепторрегулируемых Са-каналов (рецепторы Д-2). Все биол. эффекты дофамина связаны с его участием в регуляции фос-форилирования внутриклеточных белков. Биосинтез дофамина осуществляется из фенилаланина через тирозин (II) по схеме:
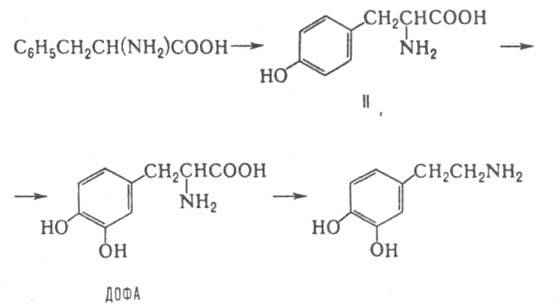
Лимитирующая стадия - синтез дигидроксифенилаланина (ДОФА), к-рая катализируется ферментом тирозингидроксилазой. Последняя активируется Са2+, поэтому факторы, повышающие концентрацию Са2+ в аксоплазме (часть цитоплазмы нейрона, входящая в состав аксона), ускоряют синтез дофамина. Стадия превращения ДОФА в дофамин катализируется декарбоксилазой ароматич. L-аминокислот. Дофамин накапливается гл. обр. в разл. отделах центр, нервной системы. В аксонах симпатич. нервных волокон и в мозговом слое надпочечников дофамин - промежут. метаболит. В этих тканях он при участии b-гидроксилазы гидроксилируется до L-норадреналина. Последний в мозговом слое надпочечников при участии фениламиноэтанол-N-метилтрансферазы и источника метильных групп S-аденозилметионина превращ. в L-адреналин. Нервные импульсы, идущие по симпатич. волокнам, приводят к выбросу L-норадреналина из пресинаптич. мембраны этих волокон и рецепции нейромедиатора на постсинаптич. мембране. В мозговом слое надпочечников секреция этих К. вызывается путем возбуждения, приходящего по чревному нерву. При этом в кровь поступают преим. L-адреналин и малые кол-ва L-норадреналина. Рецепторы этих К. есть практически во всех тканях человека и животных. L-Норадреналин взаимод. преим. с a-, a L-адреналин - с a- и b-адренергич. рецепторами. Через a-рецепторы (активируют фосфоинозитидный обмен) осуществляются вазоконстрикторные эффекты К. (приводят к сужению кровеносных сосудов), стимулируется гликогенолиз, секреция адренокортикотропина, инсулина, ренина и др. физиологически активных в-в. Через b-рецепторы (активируют аденилатциклазу) К. уменьшают периферич. сопротивление сосудов, стимулируют гликогенолиз, липолиз и белковый обмен, повышают частоту и силу сердечных сокращений. Активация синтеза К.-типичная р-ция на стресс (напр., у животных концентрация адреналина в крови под влиянием страха может возрастать в 100 раз за неск. с); их роль заключается в мобилизации организма на осуществление активной мозговой и мышечной деятельности. Важнейшую роль К. играют также в поддержании гомеостаза (постоянства состава внутр. среды и устойчивости осн. физиол. ф-ций) и адаптации организма. Удаление К. из синаптич. щели происходит путем обратного захвата в пресинаптич. окончания. Этот активный транспорт происходит с использованием энергии градиента Na+ на мембране. Часть К. диффундирует из синапса в межклеточное пространство и затем поступает в кровь. Инактивируются К. путем окислит, дезаминирования, катализируемого катехол-О-метилтрансферазой. Метаболиты К. удаляются из организма почками, поэтому их содержание в моче может указывать на активность симпатич. нервной системы и мозгового слоя надпочечников. Нарушения в метаболизме, секреции и обратном
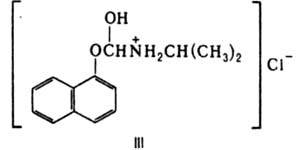
захвате К.-причина мн. патологич. процессов. Препараты, воздействующие на обмен К. и их рецепцию (напр., пропранолол, ф-ла III), широко используют для лечения ряда заболеваний человека. Лит.: Манухин Б. Н., Физиология адрснорсцепторов, М., 1968; Андреев С., Кобкова И., Роль катехоламинов в здоровом и больном организме, М., 1970; Ткачук В. А., Введение в молекулярную эндокринологию, М., 1983; Комиссаров И. В., Механизмы химической чувствительности синаптических мембран. К., 1986, с. 79-91. В. А. Ткачук.