


ВИТАМИН D (кальциферолы), группа прир.
соед., обладающих антирахитич. действием. Важнейшие: эргокальциферол [витамин
D2; рис. 1; ф-ла III, R = —СН(СН3)СН=СНСН(СН3)СН(СН3)2]
- мол. м. 396,7, т. пл. 115-118°С;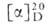 от +103° до 4-108° (абс. этанол); и холекальциферол [витамин D3;
рис. I, III, R = —СН(СНз)(СН2)3СН(СН3)2]
- мол. м. 384,6, т. пл. 84-86 °С;
от +103° до 4-108° (абс. этанол); и холекальциферол [витамин D3;
рис. I, III, R = —СН(СНз)(СН2)3СН(СН3)2]
- мол. м. 384,6, т. пл. 84-86 °С;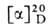 от +105° до +112° (абс. этанол). В-во, к-рое ранее называли витамином D1,
- смесь эргокальциферола и др. стеринов.
от +105° до +112° (абс. этанол). В-во, к-рое ранее называли витамином D1,
- смесь эргокальциферола и др. стеринов.
Витамины D2 и D3 - бесцв. соед., не раств. в воде,
хорошо раств. в орг. р-рителях; обладают характерным максимумом поглощения
при 265 нм (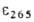 1,8*104); чувствительны к действию света и кислорода воздуха,
особенно при нагревании.
1,8*104); чувствительны к действию света и кислорода воздуха,
особенно при нагревании.
Провитамин холекальциферола-7-дегидрохолестерин [I, R = СН(СН3)(СН2)3СН(СН3)2;
содержится в коже человека] ; эргокальциферола - эргостерин [I, R = CH(CH3)CH=
=СНСH(СН3)СН(СН3)2; содержится в дрожжах].
При действии УФ-света провитамины превращ. в организме в прекальциферолы,
или превитамины (ф-ла И), из к-рых образуются витамины D в результате прототропной
перегруппировки.
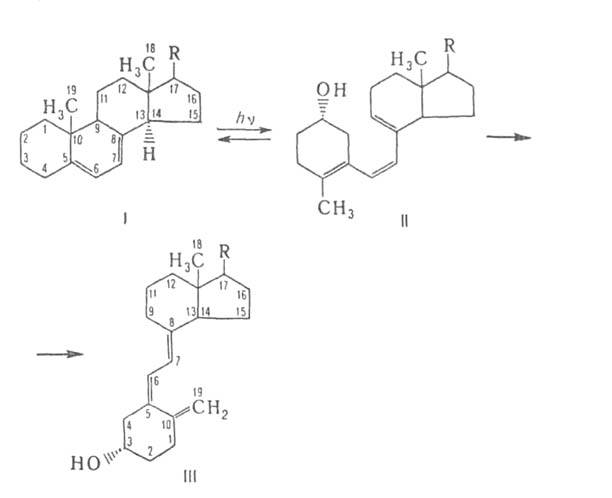
Рис. 1. Синтез кальциферолов из провитаминов.
Осн. ф-ция витамина D - поддержание в организме постоянной концентрации Са2+ и Р, что осуществляется благодаря участию витамина в регуляции всасывания этих элементов в кишечнике, мобилизации Са из скелета путем рассасывания предобразованной костной ткани и реабсорбции Са2+ и Р в почечных канальцах. Витамины D2 и D3 всасываются в тонком кишечнике и поступают в печень, где подвергаются гидроксилированию; при этом D3 (рис. 2) превращ. в 25-гидроксихолекальциферол (25-OH-D3), а D2 - в 25-гидроксиэргокальциферол (25-OH-D2). Это превращ. катализирует фермент 25-гидроксилаза. 25-гидроксикальциферолы - осн. транспортная форма витамина D в организме. В плазме крови они (как и др. формы витамина) переносятся специфич. транспортным белком - транскальциферином.
В почках из 25-OH-D3 образуются 1,25-(OH)2-D3
и 24,25-(OH)2-D3. Первое превращ. катализирует фермент
1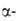 гидроксилаза,
второе -24-гидроксилаза. Аналогично из 25-OH-D2 образуются 1,25-(OH)2-D2
и 24,25-(OH)2-D2. наиб. активная форма D3,
ответственная за его ф-ции, — 1,25-(OH)2-D3 (его
биол. активность в 10 раз превышает активность.
гидроксилаза,
второе -24-гидроксилаза. Аналогично из 25-OH-D2 образуются 1,25-(OH)2-D2
и 24,25-(OH)2-D2. наиб. активная форма D3,
ответственная за его ф-ции, — 1,25-(OH)2-D3 (его
биол. активность в 10 раз превышает активность.
Важнейшие регуляторы, активирующие синтез 1,25-(OH)2-D3:
паратиреоидный гормон, эстрогены, пролактин, соматотропин и инсулин. Уменьшение
концентрации Са2+ в крови при его недостаточном поступлении
извне или интенсивной утилизации усиливает секрецию паратгормона, к-рый
активирует в почках 1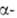 гидроксилазу,
в результате чего ускоряется синтез 1,25-(OH)2-D3.
Избыточное поступление Са2+ и Р с пищей подавляет синтез 1,25-(OH)2-D3,
т.к. при этом его предшественник 25-(OH)-D3 превращ. в почках
в 24,25-(OH)2-D3, к-рый стимулирует всасывание Са
и Р в кишечнике так же эффективно, как 1,25-(OH)2-D3,
и одновременно стимулирует остеогенез и минерализацию костной ткани.
гидроксилазу,
в результате чего ускоряется синтез 1,25-(OH)2-D3.
Избыточное поступление Са2+ и Р с пищей подавляет синтез 1,25-(OH)2-D3,
т.к. при этом его предшественник 25-(OH)-D3 превращ. в почках
в 24,25-(OH)2-D3, к-рый стимулирует всасывание Са
и Р в кишечнике так же эффективно, как 1,25-(OH)2-D3,
и одновременно стимулирует остеогенез и минерализацию костной ткани.
Осн. патоморфологич. следствие недостаточности витамина D - нарушение минерализации костной ткани. Биол. активность D2 и D3 для человека и большинства животных одинакова. Исключение-куры и нек-рые виды южноамериканских человекообразных обезьян, для к-рых D3 в неск. раз активнее, чем D2.
Биол. активность витамина D измеряется в международных (интернациональных) единицах (ME); 1 ME соответствует антирахитич. активности 0,025 мкг эрго- или холекальциферола. Содержание D2 и D3 в продуктах питания невелико, напр. в печени быка и сливочном масле соотв. 0,4 и 0,4-3,2 МЕ/г; исключение - жир печени трески и тунца, в к-рых этих витаминов содержится соотв. 50-350 и 40000-60000 МЕ/мл. Добавляя значит. кол-ва витамина D в рацион с.-х. животных, можно существенно повысить его содержание в мясе и др. продуктах.
Потребность человека в витамине D, составляющая 400 ME (10 мкг) в сут,
при достаточной и регулярной инсоляции обеспечивается фотохим. синтезом
D3 в коже. Нарушение синтеза 1,25-дигидроксикальциферолов в
почках вследствие заболеваний - причина ренальных остеодистрофий. С генетич.
дефектом синтеза этих соед. связан врожденный рахит, не поддающийся лечению
обычными дозами D2 и D3. Для профилактики и лечения
этой болезни, а также ренальных остеодистрофий наряду с препаратами 1,25-дигидроксихолекальциферола
используют также его синтетич. аналог-1 -гидроксихолекальциферол.
-гидроксихолекальциферол.
В дозах, существенно превышающих физиол. потребность, D2
и D3 высокотоксичны. Они вызывают развитие D-гипервитаминоза
с гиперкальциемией и кальцификацией внутр. органов и тканей, что ведет
к необратимым нарушениям их ф-ции и в наиб. тяжелых случаях - к летальному
исходу.
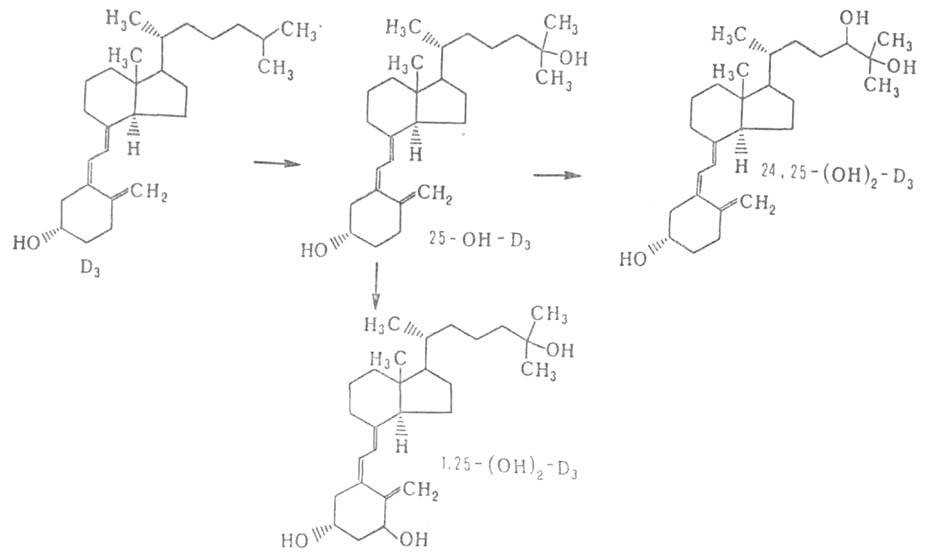
Рис. 2. Осн. пути метаболизма холекальциферола (витамина Dз).
В пром-сти эргостерин извлекают экстракцией из дрожжей; 7-дегидрохолестерин синтезируют из холестерина бромированием его эфира в положение 7 с послед. дегидробромированием. Кальциферолы получают облучением провитаминов УФ-светом в орг. р-рителе.
Лит.: Спиричев В. Б., в кн.: Витамины, М., 1974, с. 89-124; Яхимович
Р. И., Химия витаминов D, К., 1978; BekemeierH., Pfennigsdorf G., в кн.:
Fermente, Hormone, Vitamine..., 3 АиЛ., Bd 3, Tl 1, Stuttg., 1974, S. 222-327;
Norman A. W., Vitamin D the calcium homeostatic steroid hormone, N. Y.,
1979. В. Б. Спиричев.